
Never Stop Learning!
|
Never Stop Learning!
|

The beginning of a long quest
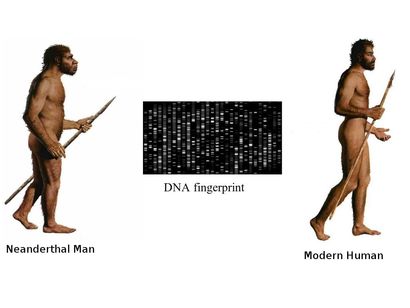
It was the year 1856 when few limestone excavators working near Düsseldorf, Germany, unveiled bones that resembled to humans and initial analysts inferred them as belonging to a deformed human, citing their oval shaped skull, with a low, receding forehead, distinct brow ridges, and bones that were unusually thick. It was only subsequent studies that revealed that the remains belonged to a previously unknown species of hominid, or early human ancestor, that was similar to our own species, Homo sapiens. In 1864, the specimen was dubbed Homo neanderthalensis, after the Neander Valley where the remains were found.Neanderthals rose to prominance around 200,000 and 250,000 years ago and ruled the hills and grasslands of europe till extiction around 30000 years ago. The exact date of their extinction had been disputed but in 2014, a team led by Thomas Higham of the University of Oxford used an improved radiocarbon dating technique on material from 40 archaeological sites to show that Neanderthals died out in Europe between 41,000 and 39,000 years ago, with the last group disappearing from southern Spain 28,000 years ago. Similarity of Neanderthals with Rhodesian Man (Homo rhodesiensis) made early investigators infer that they share similar ancestor but comparison of the DNA of Neanderthals and Homo sapiens suggests that they diverged from a common ancestor between 350,000 and 400,000 years ago, which some argue might be Homo rhodesiensis but this argument assumes that H. rhodesiensis goes back to around 600,000 years ago. However one can not rule out convergent evolutionary paths for the two hominids displaying feathres such as distinct brow ridges. Neanderthals settled in Eurasia, but not extending beyond modern day Israel. No neanderthal sites were observed in the African continent and Homo sapiens appears to have been the only human type in the Nile River Valley because of the warmer climate present in that period. Are Neanderthals really extinct? Sudden disappearnce of Neanderthals from Europe co-incides with the arrival of H. sapiens and this information prompted many scientists to suspect that the two events are closely linked, and humans contributed to the demise of their close cousins, either by outcompeting them for resources or through open conflict. The hypothesis that early humans violently replaced Neanderthals was first proposed by French palaeontologist Marcellin Boule (the first person to publish an analysis of a Neanderthal) in 1912. However according to a 2014 study by Thomas Higham and colleagues based on organic samples suggest that the two different human populations shared Europe for several thousand years. Therefore outright violent extinction seems less plausible and leads to the formation of two scenarios for Neanderthal extinction. Possible scenarios for the extinction of the Neanderthals are:
Ancient DNA to the rescue
DNA sequence analysis of the fossils can reveal an entirely new world of information to us, but recovering DNA from samples that are fossilized thousands of years ago, is a daunting task in itself making ancient DNA research far from routine. The samples are prone to degradtion and contamination by DNA from other sources, and retriving data out of the ancient material is costly and painstaking work. At a more fundamental level, it requires determining whether the necessary samples even exist and, if so, how to get access to them. The revelations An international group of Anthropologists from Max Planck Institute for Evolutionary Anthropology, Cold Spring Harbour Laboratories and Cornell University using various different methods of DNA analysis estimated an interbreeding to have happened less than 65,000 years ago, around the time that modern human populations spread across Eurasia from Africa. They reported evidences for a modern human contribution to the Neanderthal genome. Martin Kuhlwilm, co-first author of the new paper, identified the regions of the Altai Neanderthal genome sharing mutations with modern humans. They found evidences of gene flow from descendants of modern humans into the Neanderthal genome to one specific sample of Neanderthal DNA recovered from a cave in the Altai Mountains in southern Siberia, near the Russia-Mongolia border. Earlier studies have observed that DNA of modern humans contains 2.5 to 4 percent Neanderthal DNA. However studies conducted by Mendez et. al. revealed that no Neanderthal Y chromosomal DNA was ever observed in any human sample they have tested. Contemplating upon the observations they initially felt that the Neanderthal Y chromosome genes could have drifted out of the human gene pool by chance over the millennia, or there are possibilities that the Neanderthal Y chromosomes include genes that are incompatible with other human genes. Mendez, and his colleagues have found evidence supporting this idea, and they think that the two groups may have been reproductively isolated unlike thought earlier. Their study identified protein-coding differences between Neandertal and modern human Y chromosomes. Changes included potentially damaging mutations to PCDH11Y, TMSB4Y, USP9Y, and KDM5D, and three of these changes are missense mutations in genes producing male-specific minor histocompatibility (H-Y) antigens. Antigens derived from KDM5D, for example, are thought to elicit a maternal immune response during gestation. It is possible that these incompatibilities at one or more of these genes played a role in the reproductive isolation of the two groups. Thus Y-chromosomal studies have re-drawn the time-line of divergence of the two species ~4 million years ago, which according to previous estimates based on mitochondrial DNA put the divergence of the human and Neanderthal lineages at between 400,000 and 800,000 years ago. New data emerging out of GWA studies could shed further light on the evolutionary history of the two hominids. In my opinion the image could resolve better if we look into the pathogen associated and immune response genes that we might have inherited or acquired during our evolutionary journey. Reference:
0 Comments
This post is written specially keeping it consistent to the C.B.S.E curriculum for class XII. Nevertheless students from other boards can benefit from it too :)
Sexual reproduction in flowering plants (angiosperms) are carried out with the help of sexual organelles of the plant, i.e Flowers.Angiosperms: Angiosperms (Gr. Angios: Covered, Spermae: seed) are plants that have their seeds enclosed in a ovule inside the ovary of their flowers.
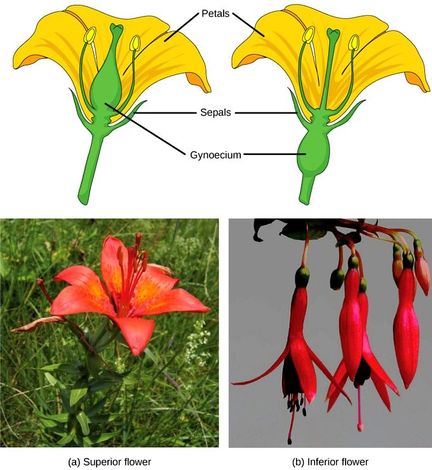
There is a huge diversity among flowers of the angiosperms but all flowers have these structures:
The ovary, which may contain one or multiple ovules, may be placed above other flower parts (referred to as superior); or it may be placed below the other flower parts (referred to as inferior).
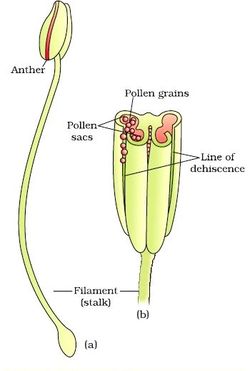
Structure of Stamen, Anther, Pollen Sac/Microsporangium and Pollen Grain in Plants! (a) The Stamen: Stamen in a flower consists of two parts, the long narrow stalk like filament and upper broader knob-like bi-lobed anther (Fig. 2 A). The proximal end of the filament is attached to the thalamus or petal of the flower. The number and length of stamens vary in different species. b) Structure of anther: A typical angiosperm anther is bilobed with each lobe having two theca, i.e they are bithecous or dithecous anther is made up of two anther lobes, which are connected by a strip of sterile part called connective. The anther is a four-sided (tetragonal) structure consisting of four elongated cavities or pollen sacs (microsporangia) the four microsporangia are located at the corners, two in each lobe. The microsporangia develop further and become pollen sacs in which pollen grains are produced.(c) Structure of microsporangium In a transverse section, a typical microsporangium appears circular in outline, consisting of two parts, microsporangial wall and sporogenous tissue. i) Microsporangial Wall: Includes the epidermis, endothecium, middle layers and the tapetum. The outer three wall layers perform the function of protection and help in dehiscence of anther to release the pollen. The innermost wall layer is the tapetum, its cells have dense cytoplasm, become large, multinucleate and are specialized in nourishing the developing pollen grains. 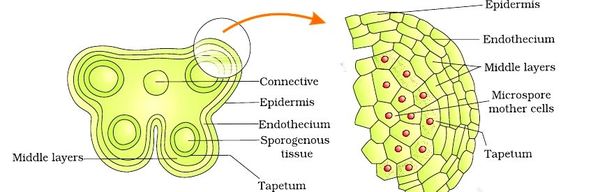
Functions of Tapetum
It fills the interior of the microsporangium, all the cells are simmilar and called sporogenous cells. Sporogenous cells devide regularly to from the diploid microspore mother cells. The microspore mother cell devides to form pollen grains. M icrosporogenesis : As the anther develops, the cells of the sporogenous tissue is capable of giving rise to a microspore tetrad. Each one is a potential pollen or microspore mother cell. The process of formation of microspores from a pollen mother cell (PMC) through meiosis is called microsporogenesis. The microspores, as they are formed, are arranged in a cluster of four cells–the microspore tetradmeiotic divisions to form microspore tetrads. 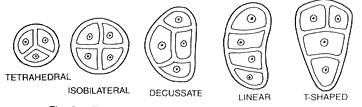
Types of microspore tetrads
As the anthers mature and dehydrate, the wall of the microspore mother cell degenerates and the microspores dissociate from each other and develop into pollen grains. Inside each microsporangium several thousands of microspores or pollen grains are formed that are released with the dehiscence of anther. 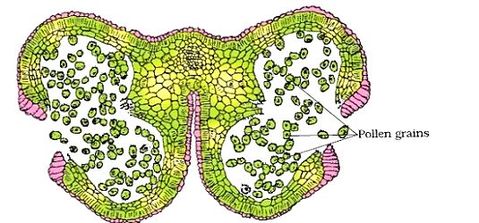
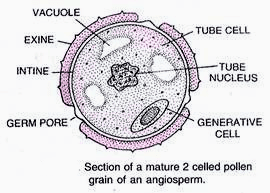
Pollen grain:
Pollen grains are male reproductive propagule or young male gametophyte which is formed in the anther and is meant for reaching the female reproductive organ through a pollinating agent. Pollen grains are generally spherical measuring about 25-50 micrometers in diameter. The pollen grains are coverd by a two-layered wall called sporoderm. The two layers of sporoderm are inner intine and outer exine. 1. Intine: It is the inner wall of the pollen grain and is a thin and continuous layer made up of cellulose and pectin. Some enzymatic proteins also occour in the intine. 2. Exine: The exine is the hard outer layer made up of sporopollenin which is one of the most resistant organic material known. It can withstand high temperatures and strong acids and alkali. No enzyme that degrades sporopollenin is so far known. Pollen grains where sporopollenin is absent can be easily identified by the presence of prominent apertures called germ pores. The exine surface may be smooth, pitted, reticulate, spiny, warty etc, the exine surface sculpting are specific for each type of pollen grain. Pollen grains are also well preserved as fossils because of the presence of sporopollenin, and thus are helpful in studying the evolutionary history of the plant. The cytoplasm of a mature pollen grain is surrounded by a plasma membrane and contains two cells, the vegetative cell and generative cell. The vegetative cell is bigger, has abundant food reserve and a large irregularly shaped nucleus. The generative cell is small and floats in the cytoplasm of the vegetative cell. It is spindle shaped with dense cytoplasm and a nucleus. In over 60 per cent of angiosperms, pollen grains of a microsporeare shed at this 2-celled stage. In the remaining species, the pollen grain generative cell divides mitotically to give rise to the two male gametes before pollen grains are shed (3-celled stage). Pollen grains of many species cause severe allergies and bronchial afflictions in some people often leading to chronic respiratory disorders – asthma, bronchitis, etc. However they are rich in neutrients and thus often consumed as food suppliment. 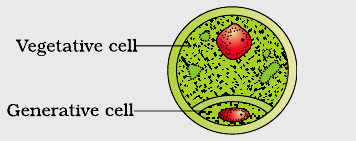
The Pistil, Megasporangium (ovule) and Embryo sac
The female reproductive parts of the flower are knwon as carpels, and are collectively called as gynoecium. Gynoecium may consist of a single pistil (monocarpellary) or may have more than one pistil (multicarpellary). When there are more than one carpel the pistils may be fused together (syncarpous) or may be free (apocarpous). Each pistil has three parts, the stigma, style and ovary. The stigma serves as a landing platform for pollen grains. The style is the elongated slender part beneath the stigma. The basal bulged part of the pistil is the ovary. Inside the ovary is the ovarian cavity (locule). The placenta is located inside the ovarian cavity. 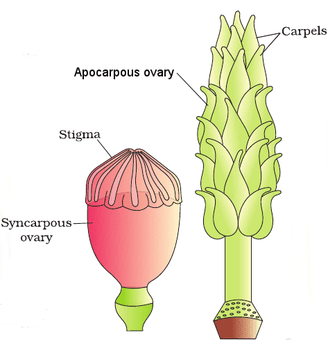
Structure of a Megasporangium (Ovule)
The ovule is a small structure attached to the placenta by means of a stalk called funicle. The body of the ovule fuses with funicle in the region called hilum. Thus, hilum represents the junction between ovule and funicle. Each ovule has one or two protective envelopes called integuments. Integuments encircle the nucellus except at the tip where a small opening called the micropyle is organised. Opposite the micropylar end, is the chalaza, representing the basal part of the ovule. The main body of the ovule is composed of parenchymatous mass called nucellus. Cells of the nucellus has abundant reserve of food. Located in the nucellus is the embryo sac or female gametophyte. An ovule generally has a single embryo sac formed from a megaspore.
Megasporogenesis
It is the process of formation of haploid megaspore from the diploid megaspore mother cell (MMC). Usually a single MMC differentiates in the micropylar region. It is a large cell containing dense cytoplasm and a prominent nucleus. The MMC undergoes meiotic division. which results in the production of four haploid megaspores, arranged generally in the form of a linear tetrad. 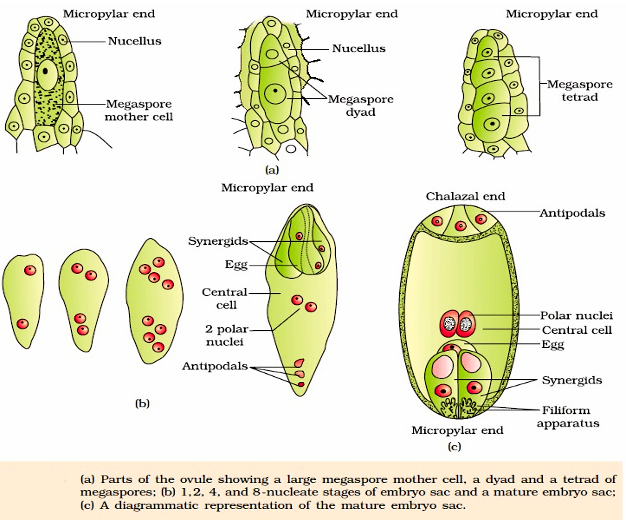
Female gametophyte or Embryo sac: Only one of the megaspores is functional while the other three degenerate. The functional megaspore develops into the female gametophyte (embryo sac).
Pollination
Pollination is the process of transferring pollen from the stamens to the stigmatic surface in angiosperms or the micropyle region of the ovule in gymnosperms. Depending on the source of pollen, pollination can be divided into three types.
Pollination mechanisms
Pollen transfer can be facilitated by the aid of abiotic (wind, water), abiotic (insects, birds, mammals).In some cases, pollen is transferred simply by gravity and the proximity of the anthers to the stigma.
Both wind and water pollinated flowers are not very colourful and do not produce nectar. 3. Zoophily It is a mode of pollination in which the biotic agents bring about pollination in flowering plants. Zoophily has several subtypes eg. Entomophily (by insects) malacophily (by snails ) chiropterophily (by Bats), ornithophilly (by birds eg. Humming bird), myrmecophily (by ants), anthrophily (by Humans). 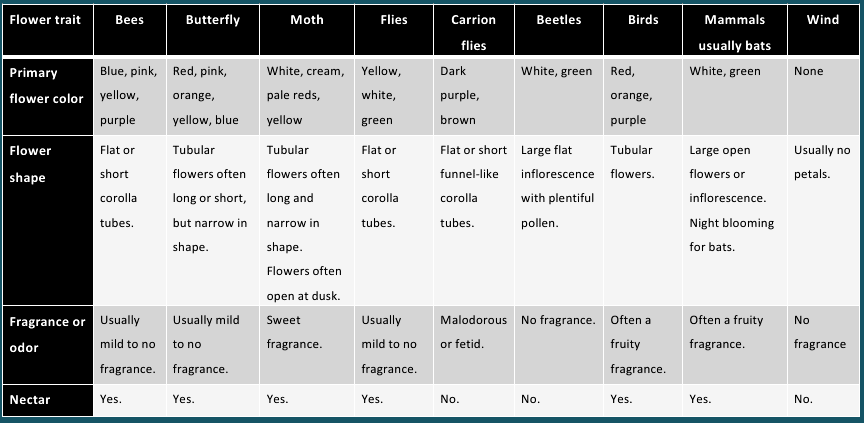
Flower traits associated with different pollination agents
Advantages and Disadvantages of Cross Pollination Advantages 1. A number of plants are self-sterile, that is, the pollen grains cannot complete growth on the stigma of the same flower due to mutual inhibition or incompatibility, e.g., many crucifers, solanaceous plants. Several plants are pre-potent, that is, pollen grains of another flower germinate more readily and rapidly over the stigma than the pollen grains of the same flower, e.g., Grape, Apple. Such plants of economic interest give higher yield only if their biotic pollinators like bees are available along-with plants of different varieties or descent 2. Cross pollination introduces genetic re-combinations and hence variations in the progeny. 3. Cross pollination increases the adaptability of the offspring towards changes in the environment. 4. It makes the organisms better fitted in the struggle for existence. 5. The plants produced through cross pollination are more resistant to diseases. 6. The seeds produced are usually larger and the offspring have characters better than the parents due to the phenomenon of hybrid vigour. 7. New and more useful varieties can be produced through cross pollination. 8. The defective characters of the race are eliminated and replaced by better characters. 9. Yield never falls below an average minimum. Disadvantages 1. It is highly wasteful because plants have to produce a larger number of pollen grains and other accessory structures in order to suit the various pollinating agencies. 2. A factor of chance is always involved in cross .pollination. 3. It is less economical. 4. Some undesirable characters may creep in the race. 5. The very good characters of the race are likely to be spoiled. Outbreeding Devices As continued self-pollination result in inbreeding depression, flowering plants have developed many devices to discourage self- pollination and to encourage cross-pollination.
Artificial hybridisation is one of the major approaches of crop improvement programme. Here only the desired pollen grains are used for pollination and the stigma is protected from contamination (unwanted pollen). This is achieved by emasculation and bagging techniques. Anthers from the flower bud before the anther dehisces using a pair of forceps is necessary. This step is referred to as emasculation.
Ref: NCERT Biology for Class 12
* All Images copyright of respective owners |
AuthorHello! My name is Arunabha Banerjee, and I am the mind behind Biologiks. Leaning new things and teaching biology are my hobbies and passion, it is a continuous journey, and I welcome you all to join with me Archives
June 2024
Categories
All



|
 RSS Feed
RSS Feed
